




|
原创世界: 【Dragicland】 头衔: 记录世界的探险家
|
| ||||











 发表于 2013-9-21 15:32
|
发表于 2013-9-21 15:32
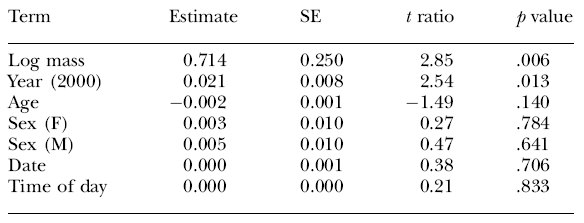
| 
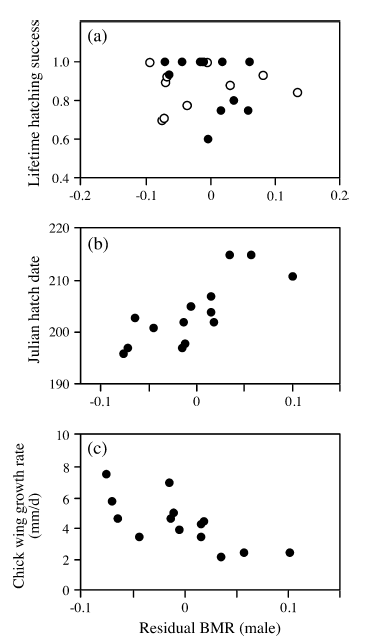
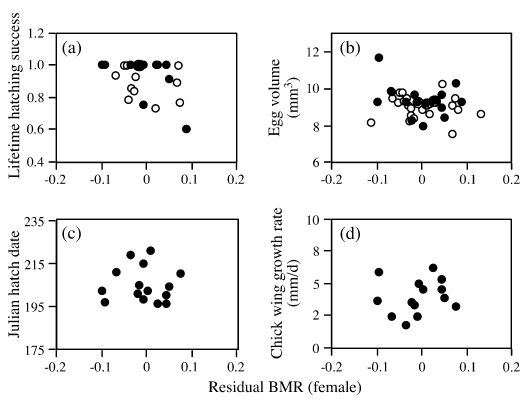
|
昵称: 喵龍
|
| |

 发表于 2013-9-28 12:01
|
发表于 2013-9-28 12:01
|
|
原创世界: 【Dragicland】 头衔: 记录世界的探险家
|
| ||||
